

Flower – A Fascinating Organ of Angiosperm
-
Flower is a sexual reproductive part of angiosperms. It develops after the completion of vegetative phase.
-
The branch of ornamental horticulture concerned with growing and marketing of ornamental flowers is called floriculture.
Parts of Flower
-
Flower has four different sets of whorls or floral members are attached to a central axis called thalamus.
-
The outermost and the first accessory whorl is of sepals and is called calyx.
-
The second accessory whorl is of petals and is called corolla.
-
Next to the corolla, is the male reproductive whorl of stamens called androecium.
-
The innermost female reproductive whorl is of carpels and is called gynoecium or pistil.
Functions of Floral Members
Calyx
-
Its main function is to protect the flower in bud condition, it is smaller than the petals in size.
-
When green, it is photosynthetic in nature.
-
When coloured, it attracts insects for pollination.
Corolla
-
It is a bright coloured attractive part of a flower and has scent to attract pollinators.
-
Sometimes the base of corolla bears some secretory glands (nectariferous gland) and secretes nectar.
-
They also provide protection to the plant.
Androecium
Its main function is the production of microspores, i.e., pollen grains containing male gametes within the anther lobe.
Gynoecium
The function of gynoecium is the production of megaspores, fruits and seeds.

Pre-fertilization structure and events
Male Reproductive Unit – Stamen, microsporangium and pollen grain
Stamen is the male reproductive unit and consists of the following two parts:
-
A long and slender stalk called filament which may be joined or free.
-
A bilobed terminal structure called anther.
Structure of Anther
-
It is composed of two anther sacs or lobes separated by a tissue called connective tissue.
-
The anther is bilobed and each lobe consists of two thecae separated by a septum.
-
The anther is a tetragonal (four-sided) structure, consisting of four microsporangia, two in each of the lobes.
-
Microsporangia develop further and get transformed into pollen sacs.
Structure of Microsporangium
-
A typical microsporangium appears circular in outline and is surrounded by four walls:
-
Epidermis: It is the outermost single layer of cell which is protective in nature.
-
Endothecium: It is the second layer with thick cells, help in dehiscence and is protective in nature.
-
Middle layer: It is the third layer composed of 1-3 layers of cells, help in dehiscence and is protective in nature.
-
Tapetum: It is the fourth and innermost layer of cell with dense cytoplasm and many nuclei. It provides nourishment to the developing pollen grains.
-
The centre of each microsporangium is filled with closely arranged similar cells called sporogenous cells.
-
At maturity, pollen sac is formed by fusion of two microsporangium in each lobe.

Microsporogenesis
-
The process of formation of microspore from a pollen mother cell by meiosis is called microsporogenesis.
-
Each cell of the sporogenous tissue in a microsporangium acts as a potential pollen mother cell (PMC) or microspore mother cell.
-
PMC undergoes meiotic divisions to form cluster of four cells called microspore tetrad.
-
On maturity, the anther dehydrates and the microspores separate from each other to form pollen grains.
Pollen Grains
-
They develop from PMC by meiotic division.
-
They represent the male gametophyte.
-
Pollen grains are generally spherical in structure.
-
They possess two prominent layered walls - outer exine and inner intine.
-
The exine is a hard layer made of sporopollenin which is one of the most resistant organic material present in nature.
-
The inner thin layer of intine is made up of cellulose and pectin.
-
The exine has an aperture where sporopollenin is absent, called germ pore.
-
The newly differentiated pollen grain has a central nucleus and dense cytoplasm.
-
The protoplast mitotically divides into two unequal cells - bigger vegetative cell which is rich in food reserve and smaller spindle-shaped generative cell with dense cytoplasm and a nucleus. This is called 2-celled stage.
-
In majority angiosperms, pollens are released in this 2-celled stage, whereas in other species, the generative cell divides into 2 male gametes and thus pollen is said to be in 3-celled stage.
Importance of pollen grains
-
They contain male gametes for sexual reproduction.
-
These are nutrient rich and thus are taken as food supplements.
-
Pollens can be stored for years in liquid nitrogen at -196 0C, to be used later in crop breeding programmes.
Female Reproductive Unit - The Pistil, Megasporangium (ovule) and embryo sac
-
The pistil or gynoecium represents the female reproductive part of a flower.
-
The gynoecium may have single pistil (called monocarpellary), more than two pistils (called multicarpellary), fused pistils (called syncarpous) or free pistils (called apocarpous).
-
A pistil has following three major parts:
-
Stigma: It receives the pollen grains. It has sticky surface and pollen grains get stuck to it during pollination.
-
Style: It is an elongated slender part beneath the stigma, that connects the stigma with ovary. It holds the stigma to receive the pollen grains. Pollen tube grows through the style and reaches the ovule.
-
Ovary: It is the basal swollen part of pistil that contains ovules and eggs.
-
Inside the ovary there is a compartment called ovarian cavity or locule separated by septum.
-
Placenta is the tissue in ovarian cavity from where ovule or megasporangium arise.
Structure of Megasporangium (Ovule)
-
The ovule is a small structure attached to the placenta by means of a stalk called funicle.
-
The junction between an ovule and a funicle is called hilum. Sometimes, the funicle extends beyond the hilum to form a ridge called raphae.
-
The ovule is surrounded by one or two protective envelopes called integuments.
-
Integument encircles the ovule entirely except at the tip, resulting in a small opening called micropyle.
-
The basal part of an ovule opposite to micropyle is called chalaza.
-
The cells with high or abundant reserve food material enclosed within integument is called nucellus.
-
The female gametophyte located within the nucellus is called an embryo sac.

Megasporogenesis
-
The process of formation of haploid megaspores from the diploid megaspore mother cell (MMC) is called megasporogenesis.
-
In the micropylar region, the nucellus contains cells with dense cytoplasm and prominent nucleus which is differentiated into a single megaspore mother cell (MMC).
-
Megaspore mother cell is diploid and undergoes meiosis.
-
MMC first divides transversely into two cells called dyad.
-
These two cells again divide transversely, as a result a linear row of four haploid cells is produced which is called megaspore tetrad or linear tetrad.
-
Out of these four megaspores, only one remains functional while the other three degenerates.
-
The one functional megaspore develops into the female gametophyte or embryo sac. This is called monosporic development.
Megagametogenesis
The formation of female gametophyte (embryo sac) is called megagametogenesis.
-
Megaspore is the first cell of the female gametophyte.
-
The megaspore increases in size and its nucleus divides mitotically into two nuclei which move apart to opposite poles. Thus, a 2-nucleate embryo sac is formed.
-
The two daughter nuclei undergo another mitotic division giving rise to the 4-nucleate stage.
-
The third mitotic division gives rise to 8-nucleate 7-celled embryo sac.
-
The central cell contains 2 nuclei known as polar nuclei.
-
The three nuclei at the micropylar region form the egg apparatus.
-
In the egg apparatus, the middle cell is the largest and is called oosphere/egg/ovum, while other two naked cells adjoining the egg cell are called synergids.
-
The three nuclei at the chalazal end are surrounded by cytoplasm and cellular wall. These are called antipodal cells.
-
The above-mentioned method of female gametophyte formation is known as normal 8-nucleate type, because 8 nuclei contribute in the formation of gametophyte. It is very common among angiosperms.

Pollination
-
The transfer of pollen grains from anther to the stigma a pistil is termed as Depending upon sources of pollen grains, pollination can be divided into .
-
Autogamy: Transfer of pollen grains from the anthers to the stigma of the same flower. E.g. pea, rice, wheat, etc.…
-
Geitonogamy: Transfer of pollen grains from the anthers to the stigma of another flower of the same plant. It is functionally cross-pollination and involving a pollinating agent. E.g. Cucurbita.
-
Xenogamy/allogamy: Transfer of pollen grains from the anthers to the stigma of a different plant of the same species. E.g. papaya, maize.
Contrivances or Devices for Self-pollination (Autogamy)
-
Cross-pollination can be prevented by exhibiting
-
Cleistogamous flowers: These are bisexual closed flowers which never open and the anthers dehisce inside these closed flowers, e.g., Commelina.
-
Homogamy: It is the condition of the maturity of anther and stigma at the same time, e.g., Catharanthus (Vinca).
-
Close association between anther and stigma, e.g., Mirabilis.
Contrivances or Devices for Cross-pollination (Xenogamy/Allogamy)
-
Self-pollination can be prevented by exhibiting
-
Unisexuality: Male and female flowers are present on different plants.
-
Dichogamy: The condition in which the stamens and stigma of a bisexual flower mature at different times.
-
Protandry: This is the condition where anthers mature earlier than the stigma and release pollens.
-
Protogyny: This is the condition where the stigma matures earlier than the anther.
-
Self-sterility or self-incompatibility: It is a genetic mechanism that prevents self-pollination.
-
Chasmogamous flowers: These are open flowers with exposed stamens and stigma which facilitate cross-pollination.
Agents of pollination
Adaptation for Water Pollination/Hydrophily
-
The form of pollination in which pollens are distributed by water is called .
-
It is very common in algae, bryophytes, pteridophytes and some angiosperms.
-
and Hydrilla are submerged freshwater hydrophytes in which hydrophily is observed.
-
In sea-grasses, female flowers are submerged in water to receive pollen grains for pollination inside water. Pollens are long, needle-like and are carried passively by water.
-
Zostera is a marine angiosperm in which hydrophily is observed.
-
In the female flower stalk is coiled to reach the water surface to receive the pollen grains carried by water currents.
-
Pollen grains are protected from wetting by the presence of a mucilaginous covering.
Adaptation for Wind Pollination/Anemophily
-
The form of pollination in which pollens are distributed by wind is called .
-
Pollen grains are light in weight, non-sticky, dry and winged.
-
Stamens are well-exposed for easy dispersal of pollen grains in the wind.
-
The stigma is sticky, large and feathery to trap pollen grains floating in the air.
-
Numerous flowers are packed together to form inflorescence.
Adaptation for Insect Pollination/Entomophily
-
The form of pollination in which insects distribute pollens is called .
-
Flowers are large, sticky and brightly coloured.
-
They have honey and nectar glands, which are highly fragrant to attract insects.
-
The pollen grain surface is sticky due to exine layer and stigma is sticky due to mucilaginous secretion.
-
The flowers offer floral rewards like nectar and pollen grains for pollination to insects.
-
In some species, floral rewards provide safe place to lay eggs,
-
The flower sometimes secretes foul odour to attract insects like flies and beetles.
Pollen - Pistil Interaction
-
All the events from pollen deposition on the stigma until the entry of the pollen tubes into the ovule are together called pollen-pistil interactions.
-
It is a dynamic process involving pollen recognition by stigma/pistil for compatible pollen.
-
Incompatible pollens or sterile pollens are rejected by the pistil and do not allow growth of pollen tube.
-
Compatible pollens are encouraged by pistil for growth and development of pollen tubes.
-
The pollen tube grows through stigma and style to reach the ovary.
-
Then it enters the ovule through micropyle reaches the synergids, guided by filiform apparatus.
Artificial Hybridisation
-
Commonly used technique in plant breeding programmes to obtain desirable characters.
-
Anthers are removed from the bisexual flowers using forceps before the anther dehisces. This step is referred as emasculation.
-
The emasculated flower is covered with a paper bag to prevent contamination from unwanted pollens. This is called bagging.
-
On attaining maturity, mature pollens from desirable plant are dusted on stigma of bagged flower and rebagged for fruit development.
Double Fertilisation/Triple Fusion
-
On reaching synergid, pollen tube releases the two male gametes into cytoplasm of synergid.
-
One of the male gamete fuses with egg nucleus to form a diploid cell called zygote. This event is called syngamy.
-
Other male gamete fuses with polar nuclei at the centre to produce a triploid primary endosperm nucleus (PEN). As this is involves the fusion of three haploid nuclei it is termed as triple fusion.
-
As syngamy and triple fusion take place simultaneously in the embryo sac, it is termed as double fertilisation.
-
The central cell after triple fusion forms primary endosperm cell (PEC) which later develops into endosperm.
-
The zygote later develops into an embryo.
Post-fertilisation: Structure and Events
-
The following events after double fertilisation are collectively called post-fertilisation events:
-
Development of endosperm from primary endosperm nucleus (PEN)
-
Development of embryo from zygote
-
Development of seeds from ovule
-
Development of fruit from ovary
Endosperm Development
-
Endosperm develops first followed by an embryo.
-
Endosperm develops from PEN.
-
The PEN undergoes successive nuclear divisions to give rise to free nuclei and this stage of endosperm development is called free nuclear endosperm.
-
Subsequently, cell wall is formed on the periphery and endosperm becomes cellular.
-
This division is followed by cytokinesis and thus endosperm becomes cellular. It is now called cellular endosperm.
Functions of Endosperm
-
The cells of endosperm tissue are triploid and filled with reserve food material to nourish the developing embryo.
-
The water of tender coconut in the centre is free-nuclear endosperm and white kernel in the outer part is the multicellular endosperm.
-
The endosperm may be completely consumed by the developing embryo, e.g., pea, beans, or it may persist in mature seed, e.g., coconut.
Embryo Development
-
Embryo develops from zygote at the micropylar end of embryo sac.
-
The nutrition for development is provided by endosperm.
-
The zygote divides to mitotically form proembryo.
-
The different stages of the developing proembryo are globular and heart-shaped embryo, which finally forms mature embryo.

Embryogeny in Dicots
-
The zygote undergoes transverse division forming a large basal cell and a small apical or terminal cell.
-
The large basal cell enlarges and undergoes transverse division to from a group of 6-10 cells called suspensor.
-
The first cell of the suspensor towards the micropylar end is called haustorium, whereas the last cell of the suspensor towards the chalazal end is called hypophysis that later develops into radicle.
-
The smaller terminal or apical cell undergoes one vertical division. The two cells formed from terminal cell divide by a transverse division thus forming four embryonal cells (quadrant stage).
-
Finally, these four cells divide vertically forming 8-celled proembryo (octate stage). Four cells at the apex give rise to plumule and another four give rise to hypocotyl except its tip.
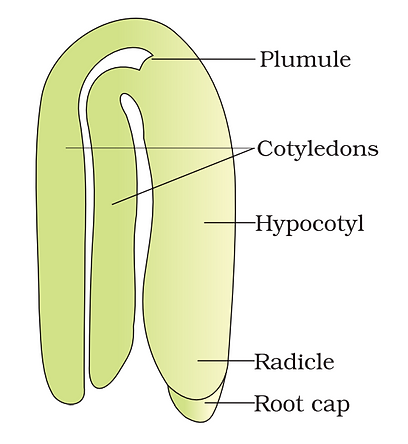
Structure of dicotyledonous embryo
-
A typical dicotyledonous embryo consists of an embryonal axis and two cotyledons.
-
The portion of embryonal axis above the level of cotyledon is called epicotyl, which develops into plumule or baby shoot (future shoot).
-
The cylindrical portion below the level of cotyledon is the hypocotyl, which develops into radicle or root tip (future root).
-
The root tip is often covered with a root cap (calyptra).
-
Mango, apple, radish, rose, etc., are some dicot plants.
Structure of monocotyledonous embryo
-
A typical monocotyledonous embryo consists of one cotyledon.
-
The cotyledon is situated on one side (lateral) of the embryonal axis and is called scutellum.
-
The radicle and root cap are situated at the lower end of embryonal axis in an undifferentiated sheath called coleorhiza.
-
The shoot apex and few leaf primordia are enclosed in a hollow foliar structure in epicotyl region called coleoptile.
-
The portion of the embryonal axis above the level of attachment of Scutellum is called epicotyl.
-
Grass, banana, bamboo, palm, etc., are some monocot plants.

Seed
-
Seed is a fertilised ovule which is the final product of sexual reproduction.
-
It consists of the following:
-
Seed coat(s): 1-2 in number, having an opening called micropyle. Outer integument forms outer seed coat (testa) and inner integument forms inner seed coat (tegmen).
-
Cotyledons: 1-2 in number and rich in reserve food material.
-
Embryonal axis
-
Sometimes in the seed, nucellus may persist. This is called perisperm, e.g., in black pepper.
-
With maturity, the water content of the seed decreases and finally enters a state of inactivity called dormancy.
Advantages of seeds
-
Seeds possess better adaptive strategies for dispersal to form a new colony.
-
The reserve food of seed supports the growth of seedling till they become nutritionally independent.
-
The hard seed coats (testa + tegmen) provide protection against injury.
-
They provide genetic recombination and variation as they are the product of sexual reproduction.
-
Seeds are stored to be consumed throughout the year, to overcome drought and natural calamities.
-
Depending upon the mobility of seeds, they are used to raise crop in favourable season.
Fruit
-
Fruit is a ripened/matured ovary. After the fertilization ovary matures into fruit and the ovarian wall develops into the wall of fruit called pericarp.
-
Fruits can be Fleshy (orange, mango etc.…,) or dry (mustard, groundnut et…,)
-
Fruits are of two types;
-
True fruit: Fruit that develops only from ovary. E.g., pea, wheat, maize et…,
-
False fruit: Fruit that develops from ovary and the thalamus or other part of flower. E.g., Apple, strawberry et…,
-
Fruits that are formed without fertilization are called parthenocarpic fruits. These do not have seeds. E.g., banana. Parthenocarpy can also be included artificially by growth hormones.
Special mechanisms of reproduction
Apomixis or agamospermy.
-
The phenomenon of asexual reproduction that imitates sexual reproduction by by formation of seed without fertilization is called apomixis or agamospermy.
-
Ways of development of apomictic seeds:
-
A diploid egg is formed without reduction division and develops into embryo without fertilisation.
-
Some cells of the nucellus, which are diploid in nature, start dividing and without fertilisation develop into embryo, e.g., citrus fruits and mango.
Polyembryony
-
The occurrence of more than one embryo in a seed is called polyembryony, e.g., orange.
-
Ways of forming polyembryonic seeds:
-
Development of cells like synergids, cells of nucellus, cells of integument, into embryo.
-
Formation of more than one embryo sac in an ovule.
Formation of more than one egg in an embryo sac.